Ⅰ. Introduction
Because of their significance in dental health, dental pulp and PDL have received considerable attention in the field of dental bioengineering [1-3]. One of the actively used methods of analysis in this regard is cDNA microarray, since it enables analysis of the expression of thousands of genes and proteins simultaneously and the comparison of the gene-expression profiles of two different samples. In case of PDL, Song et al. [4] performed a comparative gene-expression analysis between the PDL of permanent and deciduous teeth. Xie et al. [5] and Lee et al. [6] compared the microRNA profiles of human periodontal diseased and healthy gingival tissues. Han and Amar [7] examined the fibroblasts of PDL and gingiva, and de Araujo et al. [8] used cDNA microarray to study PDL cells under mechanical stress. In addition, Paakkonen et al. [9] and McLachlan et al. [10] analyzed the gene-expression patterns of healthy and carious pulp, and Paakkonen et al. [11] further investigated the differences between odontoblasts and dental pulp. Other studies have compared pairs of related dental tissues, such as the ameloblastoma and tooth germ [12], and the PDL and tooth follicle [13]. According to these researches, the candidate genes in specific function or pathologic change have been revealed.
The main functions of the PDL are tooth support, regeneration of periodontal tissues, maintenance of homeostasis, and provision of the healing process required following periodontal disease or mechanical trauma [14]. In terms of mechanical trauma, ankylosed teeth cannot be moved by orthodontic force if the PDL is absent [15]. The mechanical stress loaded onto a tooth is transferred to the PDL, the cells of which respond to that mechanical stress, regulating the absorption and formation of the bone matrix by signaling the surrounding cells [16]. The main functions of dental pulp are the formation of tertiary dentin, nerve innervation, and nutrition supply [17]. Dental pulp cells also have the ability to differentiate into odontoblasts in order to repair dental damage caused by trauma, caries, or dental erosion [18].
However, it is unknown about that the differences are made by which gene expression in spite of the functional differences of PDL and pulp. Each tissue has the specific gene expression pattern, so analysis of gene expression pattern of tissue enables to understanding the function of the tissue clearly. But no direct comparison of gene expression profiles has yet been made between the healthy normal PDL and dental pulp tissues of human permanent teeth. And, most studies were conducted by gene expression analysis using cultured cells of PDL and dental pulp. Therefore, the purpose of this study is to identify and compare the gene-expression profiles and molecular biological differences between PDL and dental pulp tissues from human normally functioning teeth.
Ⅱ. Materials and Methods
1. Sample collection
The experimental protocol was approved by the Institutional Review Board of the Yonsei University Dental Hospital (approval #2-2011-0050). Written informed consent to participate was obtained from all participants and from their next of kin, caregivers, or guardians on behalf of minors/children. The PDL samples (n=9; aged 11-19 years) and the pulp samples (n=9; aged 11-19 years) examined in this study were obtained from healthy, mature premolars extracted for orthodontic reasons from healthy persons. The extracted teeth were immediately frozen and stored in liquid nitrogen. Tissues were obtained carefully from the middle third of the PDL using sterile curettes. The teeth were subsequently crushed with a bolt cutter and the pulp was carefully obtained using sterile tweezers. The PDL and pulp were then immediately submerged in RNA-stabilizing reagent (RNAlater, Qiagen, CA, USA).
2. RNA isolation
Total RNA was extracted from the PDL and pulp samples using the RNeasy Fibrous Minikit (Qiagen) according to the manufacturer’s instructions. Prior to RNA extraction, the tissues were homogenized using a Bullet Blender Bead (Next Advanced, NY, USA). The extracted RNA was eluted in 25 μl of sterile water. RNA concentrations were determined from absorbance values obtained at a wavelength of 260 nm using a spectrophotometer (NanoDrop ND-1000, Thermo Scientific, IL, USA). The RNA samples analyzed in this study had 260/280 nm ratios of ≥1.8.
3. cDNA microarray analysis
The microarray procedure was conducted in triplicate for each sample, and using three samples from each group. Global gene expression analyses were conducted using Affymetrix GeneChip® Human Gene 1.0 ST oligonucleotide arrays (Affymetrix Inc., CA, USA). As recommended by the munafacturers protocol, 300 ng were used. Briefly, 300 ng of total RNA from each sample was converted to double-strand cDNA, as described previously [4]. Fragmented end-labeled cDNA was hybridized to the GeneChip Human Gene 1.0 ST arrays for 16 hours at 45℃ and 60 rpm, as described in the GeneChip Whole Transcript Sense Target Labeling Assay Manual (Affymetrix). After hybridization, the chips were stained and washed in a GeneChip Fluidics Station 450 (Affymetrix), and then scanned using Affymetrix Command Console software (version 1.1, Affymetrix). The raw file generated through this procedure provides expression intensity data, which were used for the next step.
4. Gene ontology analysis
Expression data were generated by Affymetrix Expression Console software version 1.1 (Affymetrix). The robust multichip average algorithm implemented in Affymetrix Expression Console software was used to normalize the data. One-way ANOVA was applied to the RMA expression values to determine whether genes were differentially expressed between the three groups. A multiple testing correction was applied to the p values of the F statistics to adjust for the false discovery rate. Genes with adjusted F-statistic p values of < 0.05 were extracted. Strongly expressed genes in PDL or pulp that exhibited differences of over 6- or 11-fold relative to the signal value of the control and each test group were selected for the further study. In order to classify the coexpression gene group, for which the expression pattern was similar, hierarchical clustering and k-means clustering were performed using MultiExperiment Viewer software, version 4.4 (www.tm4.org; Dana-Farber Cancer Institute, MA, USA). The web-based tool Database for Annotation, Visualization, and Integrated Discovery (DAVID) was used to biologically interpret the differentially expressed genes (http://david.abcc.ncifcrf.gov/home.jsp). These genes were then classified based on information on gene function from the gene ontology, Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway database (heep://david.abcc.ncifcrf.gov/home.jsp).
This microarray data set was approved by the Gene Expression Omnibus (http://www.ncbi.nlm.gov/geo/); its GEO accession number is GSE50639.
5. Quantitative polymerase chain reaction
The single-stranded cDNA required for the polymerase chain reaction (PCR) analysis was synthesized using 500 ng of extracted total RNA as a template for reverse transcription (Superscript III Reverse Transcriptase and random primer, Invitrogen, UK), as described previously [4].
The following TaqMan gene-expression assay primers were used (Applied Biosystems, CA, USA): ameloblastin (AMBN), calbindin 1 (CALB1), collagen, type XII, alpha (COL12A), dentin sialophosphoprotein (DSPP), matrix metallopeptidase (MMP)9, MMP20, secreted protein acidic and rich in cysteine (SPARC)/osteonectin 3 (SPOCK3), secreted phosphoprotein 1 (SPP1), and 18S rRNA. The results are plotted versus time, which was quantified as the cycle number. A precise quantification of the initial target was obtained by examining the amplification plots during the early log phase of product accumulation above background [the threshold cycle (Ct) number]. Ct values were subsequently used to determine ΔCt values (ΔCt=Ct of the gene minus Ct of the 18S rRNA control), and differences in Ct values were used to quantify the relative amount of PCR product, expressed as the relative change by applying the equation 2-ΔCt.
6. Immunohistochemical staining
In preparation for immunohistochemical (IHC) staining, permanent teeth were fixed in 10% buffered formalin (Sigma, MO, USA) for 1 day, decalcified with 10% EDTA (pH 7.4; Fisher Scientific, TX, USA) for 8 weeks, embedded in paraffin, and sectioned at a thickness of 3 ㎛. Specimens were subjected to IHC staining with antihuman CALB1 (rabbit polyclonal, diluted 1:400; Ab25085, Abcam, Cambrige, UK), MMP9 (rabbit polyclonal, diluted 1:800; Ab38898, Abcam), SPP1 (osteopontin, rabbit polyclonal, diluted 1:800; Ab8448, Abcam), and COL12A1 (rabbit polyclonal, diluted 1:800; Sc68862, Santa Cruz Biotechnology, Inc., CA, USA). Endogenous peroxidase activity was quenched by the addition of 3% hydrogen peroxide. Sections were incubated in 5% bovine serum albumin to block nonspecific binding. The primary antibodies were diluted to give optimal staining and the sections were incubated overnight. After incubation, EnVision+ System-HRP labeled Polymer Anti-rabbit (K4003, Dako North America Inc., CA, USA; ready to use) was applied for 20 min. Color development was performed using labeled streptavidin biotin kits (Dako) according to the manufacturers instructions. The sections were counterstained with Gills hematoxylin (Sigma). Control sections were treated in the same manner but without treatment with primary antibodies.
Ⅲ. Results
1. Gene-expression profiles of PDL and dental pulp
The total data distribution and frequency were confirmed using density and box plots and M-A plots of the standardized log intensity ratio to the average intensity. The results revealed that there was a fourfold or greater difference in expression of 347 out of 28,869 genes (1.20%) between the PDL and dental pulp tissues from permanent teeth; 83 and 264 genes were more strongly expressed in PDL and pulp tissue, respectively. Table 1 and 2 list the genes that were expressed most strongly in these two tissue types.
2. Gene ontology analysis
Gene Ontology Consortium (GO) grouping was used with the aid of DAVID to translate the microarray data into meaningful biologic functional terms and to characterize the groups of functionally related genes. The genes were classified based on information regarding gene function in gene ontology from the KEGG Pathway database. GO classes with an F-statistic p value of < 0.05 following analysis on the basis of their biologic processes and molecular functions are shown in Figure 1 and 2, respectively. In broad outlines, the GO classes of the dental pulp tissue are relatively more counted those of the PDL tissue. Especially, there were notable differences of biologic processes in regulation of cell adhesion, neurological system process, signal transduction, and ion transport, and of molecular functions in nucleotide binding, ATP binding, and ion binding.
3. Quantitative PCR
Quantitative PCR (qPCR) was performed to verify the different gene-expression levels obtained through cDNA microarray analysis. The following six genes were selected for this verification procedure: COL12, MMP9, SPP1, CALB1, SPOCK3, and DSPP. The remaining six were compared between PDL and pulp in the form of gene-expression ratios (Table 3). The pulp: PDL ratios for CALB1, SPOCK3, and DSPP were high, at 4,600, 6,800, and 15,000, respectively, while those for COL12, MMP9, and SPP1 were low, at 44, 8, and 9, respectively.
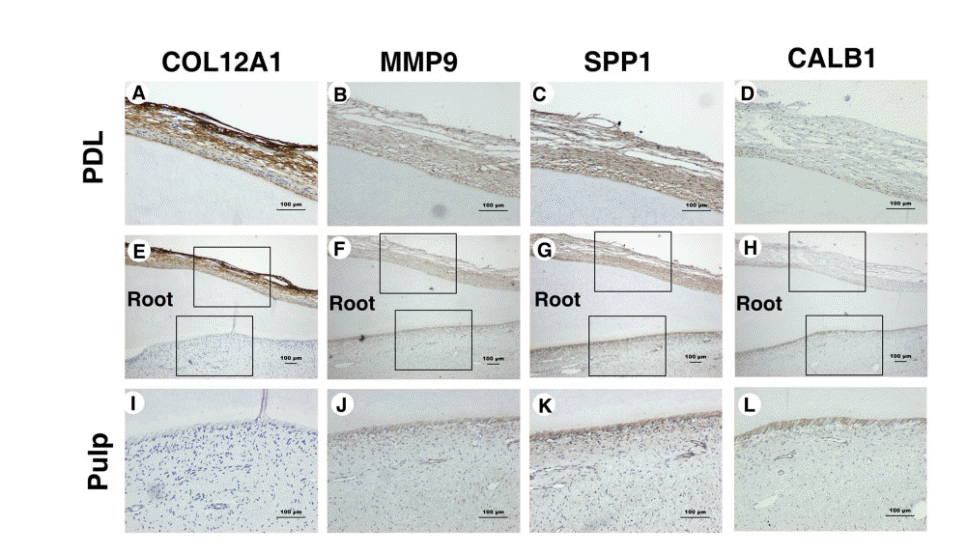
4. IHC staining
The following four proteins were the targets of the IHC study: COL12A1, MMP9, SPP1, and CALB1 (Fig. 3). COL12A1 was stained strongly in PDL tissues but was not stained in pulp tissues, while MMP9 and SPP1 were stained more strongly in all PDL tissues than in all pulp tissues, and staining was found only in the odontoblast layer of the pulp tissues. CALB1 was located mainly in the odontoblast layer of the pulp tissue.
Ⅳ. Discussion
Among the 28,869 genes that were analyzed using cDNA microarrays, 347 (1.20%) were expressed differentially by a factor of fourfold or more between PDL and pulp. According to the cDNA microarray and GO analyses, the genes that were up-regulated in the two tissue types were intimately related to their respective functions. PDL exhibited strong expression of genes related to collagen synthesis (FAP), collagen degradation (MMP3, MMP9, and MMP13), and bone development and remodeling (SSP1, BMP3, ACP5, CTSK, and PTHLH). In contrast, pulp exhibited strong expression of genes associated with calcium ions (CALB1, SCIN, and CDH12) and the mineralization and formation of enamel and dentin (SPARC/SPOCK3, PHEX, AMBN, and DSPP).
FAP alpha plays a major role in the production and turnover of extracellular matrix components, a process that is critical for wound healing and tissue remodeling. Research into FAP in dentistry is rare because of its lack of expression in normal tissues and induced expression in areas of tissue remodeling and tumor stroma [19]. MMP3, MMPR9, and MMP13 are members of the MMP family, which comprise proteolytic enzymes that mediate the degradation of extracellular matrix macromolecules and contribute not only to extracellular matrix homeostasis, but also to pathologic or therapeutic situations [20,21].
SPP1, widely known as osteopontin, has been implicated as an important factor in bone remodeling [22]. BMP3 has been extensively investigated with regard to bone formation, and reportedly down-regulates bone mineralization and density, and acts as an antagonist of osteogenic BMP [23,24]. ACP5, also known as tartrate-resistant acid phosphatase, is a glycosylated monomeric metalloenzyme that is expressed in mammals [25]. CTSK is expressed predominantly in osteoclasts; high levels of CTSK are detected in patients with bone-destructive disease. PTHLH is a member of the parathyroid hormone family that represses bone sialoprotein, osteocalcin, and mineralization in PDL cells and, potentially, in cementoblasts [26].
In the dental pulp tissues, CALB1 is an intracellular, soluble, vitamin-D-dependent calcium-binding protein and a member of the troponin C superfamily [27]. SCIN is an actin-severing protein and that is found in abundance in secretory tissues. Very little research into the gene that encodes this protein (SCIN) has been conducted in the field of dentistry, and work is needed to determine the function of SCIN in dental pulp function. CDH12 mediates calcium-dependent cell-cell adhesion. This particular cadherin appears during a critical period of neuronal development, and perhaps specifically during synaptogenesis [28]. It is thought that CDH12 up-regulation is related to the nerve innervation of dental pulp.
As expected, genes related to the mineralization and formation of enamel or dentin were strongly expressed in pulp. SPARC/SPOCK3 is a phosphorylated glycoprotein that is associated with development, tissue remodeling, and repair [29]. Phosphate-regulating endopeptidase homolog, X-linked (encoded by PHEX) is expressed by human odontoblasts aligned at the margin of the dental pulp but not in the dental pulp cells, and its expression is up-regulated during odontoblast development [30].
AMBN is found in tooth enamel, and is produced by ameloblasts during the early secretory to late maturation stages of amelogenesis. However, it has recently been shown that reparative dentin formation is associated with the sequential expression of dentin-specific factors in wounded porcine pulp [31]. DSPP is a human gene that encodes two principal proteins of the dentin extracellular matrix of the tooth.
The qPCR findings were indeed consistent with those of the cDNA microarray analysis and the IHC findings supported the cDNA microarray data and revealed the locations of the expressed genes. These data also support previous research that has shown that MMP9 may play an important role in the pathogenesis of pulpal inflammation [32], and that the SPP1 synthesized by odontoblasts is associated with the initial sites of calcification within mantle dentin [33].
Most microarray studies were about comparing the gene expression of bone marrow stem cells and stem cells derived from dental tissues. Another microarray researches were about confirming the gene expression patterns of cells under certain conditions. Although there were studies about comparative gene expression analysis of the PDL tissues in deciduous and permanent teeth and of the PDL and dental follicle tissues in humans, there is no study about comparing the relative gene expression of PDL and dental pulp tissues in humans. So gene expression analysis might help to explain the functional difference between PDL and dental pulp.
In conclusion, comparison of the PDL and the dental pulp tissues revealed genetic differences of actively formationing gene. The gene-expression profiles presented here identify candidate genes that may distinguish specific functions of PDL and dental pulp. The critical point of this research is that RNA obtained from fresh functioning dental pulp and PDL tissues of permanent teeth, showed different specific genetic activities. And in tissue regenerative therapy, it might help to find the key regenerative factors in PDL and pulp regeneration if further studies are performed about genes with different expression pattern in two types of tissues.
Ⅴ. Conclusion
This study was conducted to identify the gene-expression profiles and their molecular biological differences of PDL and dental pulp tissues from the human permanent teeth using cDNA microarray analysis, qPCR, and immunohistochemical stain. Genes associated with collagen degradation such as MMP3, MMP9, and MMP13 and with collagen synthesis such as FAP, and with bone development and remodeling such as SPP1, BMP3, ACP5, CTSK, and PTHLH were more strongly expressed in PDL tissues of permanent teeth. They are clinically related to PDL’s functions of external force absorption and tooth supporting. In dental pulp tissues of permanent teeth, genes associated with calcium ion such as CALB1, CDH12, and SCIN and with mineralization and formation of enamel or dentin such as SPOCK3, PHEX, AMBN, and DSPP were more strongly expressed. They are clinically related to dental pulp’s functions of secondary and tertiary dentin formation. The qPCR analysis and immunohistochemical staining analysis was also coincided with the cDNA microarray assay data.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print