Ⅰ. 서 론
치아의 발달은 상피(epithelial)와 간엽(mesenchymal) 세포의 상호작용으로 형성된다. 이러한 치성 세포는 각각 구강 외배엽(oral ectoderm)과 하부 간엽 조직(underlying mesenchyme)에서 기원한다. 치아에서 상피-간엽 상호작용(epithelial-mesenchymal interaction)은 순차적(sequdntial)이고 상호유도적(reciprocal)인 일련의 중요한 기전이다[1]. 그 중 치아유두세포(dental papilla cell)는 상아질모세포(odontoblast)로 분화하여 상아질을 형성하는 기능을 담당한다[2]. 상아질모세포는 상아질을 분비하는 세포로, 평생에 거쳐 전상아질(predentin)과 2차 및 보상성 상아질을 침착시킨다[3]. 또한 상아질모세포 특이 유전자로 알려진 Alkaline phosphatase (ALP), Dentin Matrix Protein 1 (DMP-1), Dentin sialophosphoprotein (DSPP), Osteocalcin (OCN) 그리고 Osteonectin (ONT) 등이 상아질모세포 전구세포(preodontoblast)에서 상아질모세포로 분화되는 과정에서 발현된다[3-5].
과잉치는 정상 치열에서 과잉의 개수를 보이며, 유병률은 0.15 - 1.9%이다. 남아에서 자주 발견되고, 구개측에서 단독으로 역위된 형태가 가장 많이 발견된다[6]. 인접치 맹출 지연, 부정교합 그리고 함치성 낭 등의 합병증을 초래할 수 있어, 만 10세 이전의 비교적 조기에 발거하는 것이 추천된다[7].
인간 줄기세포(human stem cells)는 여러 종류의 신체 조직으로 분화할 수 있는 능력을 가진 세포 즉, 미분화 세포이다. 치수, 탈락한 유치, 치주인대, 치낭 그리고 치유두와 같이 치아 및 주변조직에서 얻을 수 있다[8]. 성체 줄기세포는 법랑질을 제외한 상아질, 치주인대, 백악질 그리고 치수조직으로 분화가 가능하여[9], 그 연구 가치가 크다고 할 수 있다. Gronthos 등[10]에 의하면, 사람 치수 세포에서 자가 재생 능력(self-renewal capacity) 및 지방세포나 신경세포로의 분화 능력(differentiation capacity), 그리고 클론 형성 효율(clonogenic efficiency)이 관찰되어 줄기세포와 같은 특성을 가지고 있음을 보고하였다. 또한 Seo 등[11]에 의하면, 치주조직의 항상성을 유지하는 기능을 담당하는 사람 치주인대 세포에서 백악모세포-유사 세포(cementoblast-like cells), 지방세포, 그리고 교원질-형성 세포(collagen-forming cells)로 분화하는 것을 밝혀, 이 역시 줄기세포와 같은 특성으로서 조직 재생의 가능성을 보고하였다.
제3대구치 혹은 유치의 치수 또는 치주인대 세포를 비교한 연구가 있으나 과잉치에 대해 연구한 논문이 부족한 실정이다. 저자는 정량적 실시간 중합효소 연쇄 반응법(quantitative real-time polymerase chain reaction, qRT-PCR)을 이용하여 발거된 과잉치의 치수 및 치주인대 줄기세포의 유전자 발현 특성에 대해 비교 연구해보고자 한다.
Ⅱ. 연구 대상 및 방법
1. 과잉치 치수 세포 및 치주인대 세포의 채취
이 연구는 2016년 상악 매복 과잉치를 주소로 단국대학교 치과대학 부속치과병원에 내원한 전신질환 및 의과적 병력이 없는 만 6세의 남아 2명을 대상으로 하였다. 과잉치의 치수 및 치주인대 조직의 채취는 환자 및 보호자의 서면동의 하에 이루어졌다. 정중부에 역위된 과잉치는 숙련된 술자의 계획 하에 단국대학교 치과병원 소아치과에서 발거하였다. 과잉치는 발거된 즉시 α-minimum essential medium (α-MEM; Gibco BRL, Grand Island, NY, USA)에 보관하여 실험실로 옮겨 세포의 손실을 최소화하였다. 발거 당일 치수 조직과 치주인대 조직으로부터 세포를 채취하여 10% fetal bovine serum (GIBCO, Life Technologies, Carlsbad, Calif., USA)을 포함하는 α-MEM media에서 일차 배양하였다.
2. 세포의 분리 및 배양
각 세포의 추출은 멸균된 공간에서 시행하였다. 치수 세포를 추출하기 위해 과잉치의 백악법랑경계 하방에서 멸균된 생리식염수 주수 하에 고속 절삭기구를 사용하여 치수가 노출되지 않을 정도까지 삭제하였다. 이후 치수를 노출시키고, 멸균된 파일을 이용하여 치수 세포를 채취하였다. 치주인대세포를 채취하기 위해 치근 중앙 1/3부위의 치은접합부위(gingival attachment)와 치근단에서 최소 2 mm 떨어진 부분에서 No. 15 blade를 이용하여 조직을 분리했다[12]. 얻어진 조직은 1 mm3의 크기로 잘게 자른 후 Enzymatic digestion 방법을 적용하여 3 mg/mL의 Type I collagenase (Sigma-Aldrich Co., St. Louis, MO, USA) 와 4 mg/mL 의 Dispase (Sigma-Aldrich Co., St. Louis, MO, USA)를 넣고 37℃, Shaking incubator에 1시간 동안 효소와 반응이 잘 일어나도록 두었다. 이후 하나의 세포를 얻기 위해 70 μm Falcon strainer (CORNING Inc., NY, USA)로 거른 후, 필터를 통과한 세포를 37℃, 5% CO2의 습윤 항온기에서 4 - 7일간 배양하였다. 배지는 48시간 후부터 2 - 3일에 한 번씩 바꾸었으며, 배지를 교체할 때 배양 용기에 부착되지 않은 부유물들을 씻어냈다. 부착된 세포가 배지의 80% 이상을 차지할 때 Trypsin-EDTA (CORNING Inc., NY, USA)로 세포를 분리시켰다. 과잉치의 치수 및 치주 조직으로부터 얻은 세포는 각각 supernumerary dental pulp stem cells (sDPSCs), supernumerary periodontal ligament stem cells (sPDLSCs)로 명명하였다. 이후 1/4씩 나누어 동일한 방법으로 3계대까지 배양하였다.
3. 정량적 실시간 중합효소 연쇄반응을 통한 유전자 발현 관찰
Easy-spin total RNA extraction Kit (iNtRON Biotechnology, Gyeonggi-do, Korea)를 사용하여 각 세포에서 Total RNA를 추출하였고, Nanodrop ND-2000® (Thermo Scientific, Waltham, MA, USA) 분광광도계로 흡광도를 측정한 후 RNA의 정량을 시행하였다. 이후 주형 cDNA를 합성하기 위하여 qPCR RT Master Mix kit (TOYOBO Co., Osaka, Japan)을 이용하여 RNA의 역전사 반응을 유도하였다.
qRT-PCR 연구를 위해, 유전자 증폭에 필요한 polymerase가 적절히 섞여있는 syber green master mix (Bio-Rad Labratories, Hercules, CA, USA)를 사용하였다. 분석을 위한 Primer는 alkaline phosphate (ALP), osteocalcin (OCN), osteonectin (ONT), dentin matrix acidic phosphoprotein 1 (DMP-1), dentin sialophosphoprotein (DSPP)이다. 외부조건에 잘 변하지 않고 대부분의 세포에서 고르게 발현되는 glyceraldehyde 3-phosphate dehydrogenase (GAPDH)을 기준으로 유전자의 상대적 발현량을 비교하였다.
중합효소 연쇄반응(polymerase chain reaction, PCR)은 StepOnePlus™ Real-time PCR (AB Applied Biosystems by life Technology, Waltham, MA, USA)을 이용하여 분석하였다. 초기 변성(denaturation)으로서 95℃에서 20초, 소둔(annealing)은 각 primer의 적정 온도에서 1분, 총 40 cycle로 진행하였다. 또한 융해곡선(melting curve) 분석을 위해 65℃에서 95℃까지 해리(dissociation) 과정을 함께 진행하였다. 융해곡선이 one peak로서 단 하나의 PCR product가 생성되는지 검증하였다. 사용한 primer의 sequence와 소둔 온도는 Table 1에 표기하였다.
Ⅲ. 연구 성적
실시간 중합효소 연쇄반응(Real-time PCR)을 통해 얻은 ALP, OCN, ONT, DMP-1, DSPP와 Housekeeping Gene으로 사용한 GAPDH와의 threshold cycle (Ct)값을 얻었다. 이를 통해 △Ct (target gene - housekeeping gene)를 구하고 그 값을 이용해 유전자의 상대적인 발현량을 알아보았다.
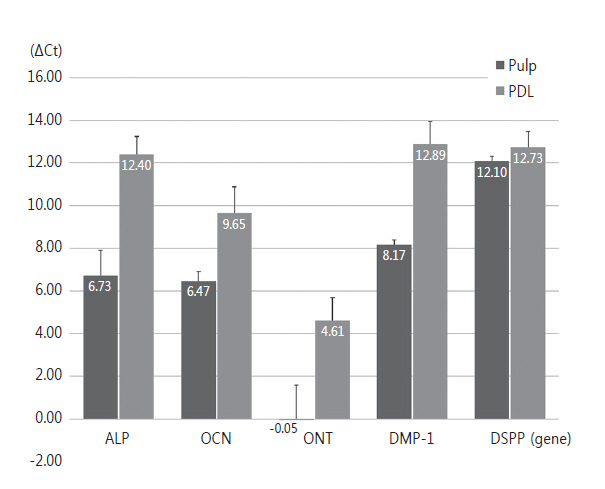
전체적인 △Ct (target gene - housekeeping gene)값은 Table 2에 나타내었다. 이 값들을 그래프로 표현한 것은 Fig. 1이다. △Ct (target gene - housekeeping gene)이 작게 나타날수록 처음 사용된 시편에 포함된 유전자의 숫자는 많다는 것을 의미한다. 따라서 과잉치의 치수 세포에서는 ONT가 가장 많고, DSPP가 가장 적게 존재하였다. 또한 과잉치의 치주인대 세포에서는 ONT가 가장 많고, DMP-1이 가장 적게 존재하였다.
치수 세포보다 치주인대 세포에서 △Ct 값의 변화는 모두 증가하였다. Mann-Whitney U test로 통계적 유의차를 검증해본 결과 모두 유의한 차이를 보이지 않았다.
Table 3는 치수 세포와 치주인대 세포에서의 상대적 유전자 발현량을 순서대로 나타내었다. 치수세포에서는 ONT, OCN, ALP, DMP-1, DSPP 순서로 많이 발현하였다. 치주인대 세포에서는 ONT, OCN, ALP, DSPP, DMP-1 순서로 많이 발현하였다. 치수세포와 치주인대 세포를 비교할 때, ONT, OCN, ALP의 순서는 변화가 없었지만, DMP-1과 DSPP는 순서가 바뀌었다.
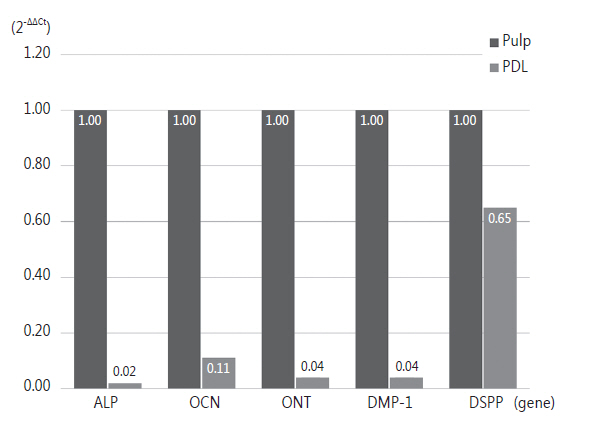
Table 4는 치수 세포와 치주인대 세포에서 △Ct (target gene - housekeeping gene)값의 차이를 2의 배수 값으로 환산한 값으로 상대적 정량[13]을 시행하였다. 이 값들을 그래프로 표현한 것은 Fig. 2이다. 치수 세포에 대한 치주인대 세포의 발현량을 비교한 경우, 치수 세포보다 치주인대 세포에서 ALP, OCN, ONT, DMP-1, DSPP 모두 발현량이 감소하였다. Mann-Whitney U test로 통계적 유의차를 검증해본 결과 모두 유의한 차이를 보이지 않았다.
Table 5은 치수 세포에 대한 치주인대 세포의 발현량을 증가한 순서대로 나열하였다. 이는 DSPP, OCN, ONT, DMP-1, ALP 순서였고, 치주인대 세포에서 치수 세포에 비해 모든 유전자의 발현이 감소하였다.
Ⅳ. 총괄 및 고찰
이 연구의 목적은 정량적 실시간 중합효소 연쇄반응법(Quantitative Real-time Polymerase Chain Reaction, qRT-PCR)을 이용하여 발거된 과잉치 치수(supernumerary dental pulp stem cells; sDPSCs) 및 치주인대 줄기세포(supernumerary periodontal ligament stem cells; sPDLSCs)의 상아질모세포 특이 유전자로 알려진 ALP, OCN, ONT, DMP-1, 그리고 DSPP의 발현 특성에 대해 알아보는 것이었다.
정량적 실시간 중합효소 연쇄반응은 분자 생화학에서 유전자 발현에 대한 이해를 증진시킬 수 있는 방법이다. 이는 목표 DNA 분자의 증폭과 양의 측정을 동시에 시행해서 증폭된 DNA가 실시간으로 측정되며[14], 이를 통해 Ct값을 얻는다. Ct값은 증폭된 target의 양이 고정된 역치(threshold)에 도달하는 부분 사이의 주기(cycle) 수를 의미하며, target에서 reference를 빼서 구한다[13]. 이 연구에서는 reference로 GAPDH를 사용하였다. 따라서 Ct값이 작을수록 시편에 포함된 유전자의 숫자가 많다는 것을 의미한다. 또한 본 연구에서는 발현된 유전자 양을 해석하기 위한 방법으로 상대적 정량법을 사용하였다. Livak 등[13]이 제안한 2-△△Ct 방법을 이용하여 분석하였다.
과잉치는 병인이 정확히 밝혀지지 않았으나 유전 혹은 환경 요인으로 복합적이다. 일반적으로 초기 혼합치열기에 맹출하지 않은 상태로 발견되는 경우가 대부분이다[15]. 과잉치를 적절한 시기에 발거하지 않았을 때 정중이개 혹은 영구치 맹출과 배열에 문제가 생길 수 있어, 수술적 혹은 교정적 개입이 필요한 경우가 있다[16]. Shih 등[15]은 5세 이전에 과잉치를 발거하는 경우, 합병증과 교정치료 요구를 줄일 수 있다고 제안했다. 비교적 이른 시기에 과잉치 발거를 시행하는 경우, 줄기세포의 공여부로서 어린 세포를 얻을 가능성이 높다고 예측해볼 수 있다.
상아질모세포는 외배엽성 중간엽 세포(ectomesenchymal cells)의 배아 세포(embryonic connective cells)에서 기원하고, 중요한 기능으로서 상아질을 형성하고, 반응성 또는 보상성 상아질의 새로운 층을 침착시키며, 감각을 담당한다[17]. Couve 등[18]은 상아질모세포 전구세포(pre-odontoblasts)가 미성숙하고 분극화(polarized)된 상태로 치근의 끝에서부터 자라기 시작하여 연속적인 단계를 거쳐 분화한다고 밝혔다. 이후 분화되어 더욱 활동적으로 분비(secretory)하는 시기가 되면 1차 상아질(primary dentin)이 형성된다. 치관부 1차 상아질의 형성이 완료되면, 비교적 분화 후기의 나이 든(aged) 상아질모세포가 이행 단계(transitional stage)를 거친다. 치아 맹출 이후 교합을 형성하면, 2차 상아질의 형성이 시작되어 평생동안 이루어진다[19]. 상아질모세포가 분화하는 과정에서 상아질모세포 전구세포는 주로 ALP, OCN 그리고 ONT를 형성하고[16], 분화한 상아질모세포는 bone sialoprotein (BSP), DMP-1, DSPP, 그리고 osteopontin (OPN)을 형성한다[4,20].
이 연구에서 사용된 primer인 ONT, OCN, 그리고 ALP는 골 형성 표지자(osteogenic marker)로, DMP-1과 DSPP는 상아질 형성 표지자(dentinogenic marker)로 설정하였다. ONT는 수산화인회석(hydroxyapatite)과 교원질(collagen)에 선택적으로 작용하는 골-특이적(bone-specific) 단백질이다[21]. 골모세포와 상아질모세포에서 분비되며, 분화 초기에 발현된다[5]. 본 연구에서 ONT는 치수 및 치주인대 세포에서 가장 많이 발현되었다. 이는 비교적 초기 계대인 3계대를 사용하였기 때문으로 예측되며, ONT가 비교적 분화 초기에 발현된다는 기존 연구와 일치한다. OCN은 치수 손상에 대한 반응으로 발현되는 보상성(reparative) 분자로서, 3차 상아질에서도 발견된다. 이는 골모세포의 표지자이며, 상아질모세포의 분화 후기에 나타난다[22]. 본 연구에서 OCN은 치수 및 치주인대 세포에서 두번째로 많이 발현되었다. 증가량을 비교해보면, ONT에서 치수 세포는 91.77배, 치주인대 세포는 32.90배 증가한 것으로, 가장 많이 증가하였다. ONT와 OCN은 각각 첫번째와 두번째로 많이 발현되었고 순위에서는 큰 차이가 없지만, 유전자 양은 상당한 차이가 있는 것을 알 수 있었다. ALP는 무세포성 치근 백악질의 형성과 상아질의 무기질 층을 형성하는 기능을 담당하는 효소이다[23]. 또한 증령에 따라 활성도가 감소하여 치수와 상아질 회복(repair)에 영향을 미치는 것으로 알려져 있다[24]. Park 등[25]의 과잉치 치주인대 줄기세포의 특성 연구에서 ALP는 분화 초기인 1 - 2주에 발현이 증가하다가 다시 감소하는 경향을 보였다. 본 연구에서 ALP는 치수 및 치주인대 세포에서 세번째로 많이 발현되어 중간 정도의 발현량을 보였다. 하지만 증가량을 비교해보면, OCN에서 치수 세포는 1.20배, 치주인대 세포는 6.73배 증가를 보였기 때문에 앞서 비교한 ONT와 OCN에 비해서는 비교적 차이가 크지 않았다.
DMP-1은 비교원성 단백질로서, 골과 치아의 석회화에 중요한 기능을 담당한다[26]. Alessandra 등[27]은 DMP-1이 전상아질층에 가까운 2차 상아질에서 많이 관찰된다고 보고했다. 그러나 Couve 등[3]은 DMP-1의 발현이 1차 상아질을 형성하는 상아질모세포 분화 초기에 많이 관찰되며, 성숙한 상아질모세포에서는 적게 관찰된다는 반대의 결과를 보고했다. 본 연구에서 DMP-1은 치수 및 치주인대 세포에서 각각 4번째, 5번째로 비교적 적은 양이 발현되었기 때문에 전자의 결과와 일치하였다. DSPP는 DMP-1과 마찬가지인 비교원성 단백질로, 골과 치아와 같은 경조직의 대표적인 상아질 특이 단백질이다[28]. Alessandra 등[27]은 DSPP가 전상아질층에서 많이 관찰되었다고 보고했다. 그러나 Papagerakis 등[5]은 DSPP가 완전히 분극화된 상아질에서만 발견되어, 분화의 비교적 후기에 관찰된다고 보고하였다. 본 연구에서 DSPP는 치수 및 치주인대 세포에서 각각 5번째, 4번째로 비교적 적은 양이 발현되었기 때문에 후자의 결과와 일치하였다.
2004년 Kamata 등[29]의 제3대구치 치수 및 치주인대 세포의 cell line을 이용한 연구에서, 치수 세포에서 alkaline phosphatase (ALP), osteopontin (OPN), osteocalcin (OCN), 그리고 dentin sialophosphoprotein (DSPP)이 발현되었다고 보고하였다. 그러나 치주인대 세포에서는 ALP, OPN, OCN은 나타나는 반면, DSPP는 발현되지 않았다고 보고하였다. 하지만 본 연구에서는 치주인대 세포에서도 DSPP가 4번째로 많이 발현되었고, 상대적 정량법의 결과에서는 치수 세포에 비해 치주인대 세포에서 상대적으로 가장 많이 발현되었다. 이는 조직 채취 과정에서 치주인대 조직 외에 치근 주위 조직이 함께 채취되었을 수 있으나, 두 표본 모두에서 발현되었기 때문에 그 가능성이 낮을 것으로 사료된다. 또한 제3대구치와 과잉치 세포의 차이에서 기인했을 가능성도 있으므로, 앞서 언급한 사항들을 보완하는 추가적인 연구가 필요할 것으로 사료된다.
제3대구치를 이용한 2009년 Hyoun 등[30]의 연구에서는 치수세포에서 2배 이상 증가된 발현을 나타내는 유전자가 치주인대 세포에서 2배 이상 증가된 발현을 나타내는 유전자 수보다 훨씬 많이 나타났다고 보고하였다. 또한 이 연구와 동일한 유전자를 사용하지는 않았지만, 상아모세포에서 많이 발현된다고 밝혀진 유전자들 중 몇 개가 특징적으로 관찰되었다고 보고하였다. 이 연구에서 유전자 발현량의 순위와 각 유전자의 분화 시기를 고려해보면, 과잉치 치수 및 치주인대 세포 모두 상아질모세포의 특성을 보인다고 예측해볼 수 있었다. 또한 이 연구에서 사용된 상아질모세포 특이 유전자 모두에서 치주인대 세포보다 치수 세포에서 유전자의 상대적 발현량이 높은 것으로 관찰되어 일치하는 결과를 보였다. 반면 치주인대 세포에서는, 치주조직의 항상성을 유지하는 교원질 합성과 분해에 관여하는 특징적인 유전자 몇 개가 관찰되었다. 그 결과 치수 세포에는 상아질 형성에 관여하는 특징적인 유전자가, 치주인대 세포에는 교원질 합성에 관여하는 특징적인 유전자가 상대적으로 높게 발현된다고 보고하였다. 따라서 추후 과잉치 치주인대 세포에서의 교원질 관련 유전자 발현에 대한 추가 연구가 필요할 것으로 보인다.
또한 이 논문의 한계로서, 사용된 표본의 수가 2개로 매우 적었기 때문에, 추후 제3대구치와 유치 및 과잉치를 비교할 수 있는 더 많은 수의 시편에 대한 추가 연구가 필요할 것으로 판단된다.
Ⅴ. 결 론
이 연구는 과잉치 치수 세포 및 치주인대 세포의 차이를 알아보고자 qRT-PCR을 사용하여 유전자 발현 정도를 비교하였다. 과잉치 치수 및 치주인대 세포 모두 상아질모세포의 특성을 보이는 것을 확인할 수 있었다.
과잉치 치수 세포에서는 ONT가 가장 많이 발현되었으며, 다음으로 OCN, ALP, DMP-1, DSPP 순서였다. 과잉치 치주인대 세포에서 역시 ONT가 가장 많이 발현되었으며, DMP-1과 DSPP의 순서만 바뀌었다. 또한 모든 유전자는 치수 세포에서 치주인대 세포보다 유전자 발현량이 많았으며, 치수 세포에 대해 치주인대 세포에서 DSPP의 상대량이 가장 많았다.
이러한 상아질모세포의 특성을 고려해 보았을 때, 다른 조직으로의 분화 가능성이 있는 과잉치 줄기세포는 유용한 공여부로서 그 잠재력이 있음을 알 수 있었다.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print